Las neuronas en desarrollo tienen la propensión a aprender sobre la marcha
No está claro cómo contribuyen los factores genéticos y ambientales a la generación de varios subtipos de neuronas inhibitorias en el cerebro llamadas interneuronas. Un estudio en ratones proporciona una nueva visión de este proceso.
Christian Mayer Gord Fishell
El cerebro maduro contiene una enorme variedad de neuronas inhibitorias, que se proyectan localmente, conocidas como interneuronas. Cómo se genera el complemento preciso de las interneuronas del cerebro durante el desarrollo es un tema de intenso debate. En el fondo, esta pregunta tiene que ver con el dilema genes versus ambiente. Las jóvenes interneuronas 'nacen' en una región llamada subpallium y experimentan una larga migración para llegar a sus posiciones finales en la corteza cerebral, pero no está claro cuánto del destino maduro de una interneurona se debe a su identidad genética, que se establece cuando la célula deja de proliferar, y cuánto se adquiere por efecto del ambiente durante la migración. En un artículo escrito en Nature Neuroscience , Lim et al. investigaron cómo influye la migración en la identidad celular.
Los autores estudiaron las vías de migración de dos tipos de interneuronas: una caracterizada en el adulto por la expresión de la proteína somatostatina y la otra por la expresión de la proteína parvalbúmina. Ambos tipos nacen en la misma región general del subpallium. Las interneuronas de esta región alcanzan la corteza embrionaria predominantemente por dos rutas de migración: una que las lleva a través de la zona marginal por encima de la corteza; y una que transita por debajo de la corteza, a través de la zona subventricular.
Hay evidencia para apoyar los roles tanto de los genes como del ambiente en la definición de las identidades de las diferentes clases, tipos y subtipos de interneuronas en el cerebro maduro. Se cree que los programas intrínsecos de expresión génica comienzan a definir las identidades de las interneuronas en el embrión, y se desarrollan durante un largo período de tiempo. La expresión de ciertos genes permanece conservada en tipos específicos de interneuronas desde su nacimiento hasta la edad adulta, mientras que otros afectan la maduración interneuronal más transitoriamente durante el desarrollo embrionario temprano. Se cree que tales procesos intrínsecos cooperan con el entorno de la interneurona para establecer circuitos neuronales y conectividad cerebral en el adulto. Por ejemplo, después de la llegada a la corteza cerebral, la actividad neuronal afecta varios aspectos del desarrollo interneuronal.
¿La ruta tomada por una interneurona inmadura tiene un efecto sobre la identidad de la célula madura en la que se convertirá? Para descubrirlo, Lim et al.específicamente etiquetaron los precursores de las interneuronas, que expresan somatostatina o parvalbúmina en la zona marginal, con una proteína fluorescente, y observaron el desarrollo de las células. Los autores encontraron que ambas poblaciones tienden a desarrollar proyecciones complejas al final de su migración a través de la zona marginal. Estas proyecciones, llamadas axones translaminares, atraviesan diferentes capas de la corteza. Este hallazgo llevó a los investigadores a proponer que la migración a través de la zona marginal influye en el crecimiento y la ramificación de los axones a través de algún mecanismo general.
Para examinar esta idea, Lim y sus colegas realizaron experimentos de imágenes time-lapse en cortes de cerebro en cultivo. De acuerdo con su hipótesis, las interneuronas 'descendieron' hacia la corteza después de viajar a través de la zona marginal. Durante este proceso, la mayoría de estas células anclaron su axón naciente en la zona marginal como una cuerda de arrastre (Fig. 1). Por lo tanto, la ruta de migración y el desarrollo axonal parecen estar vinculados para estas células.
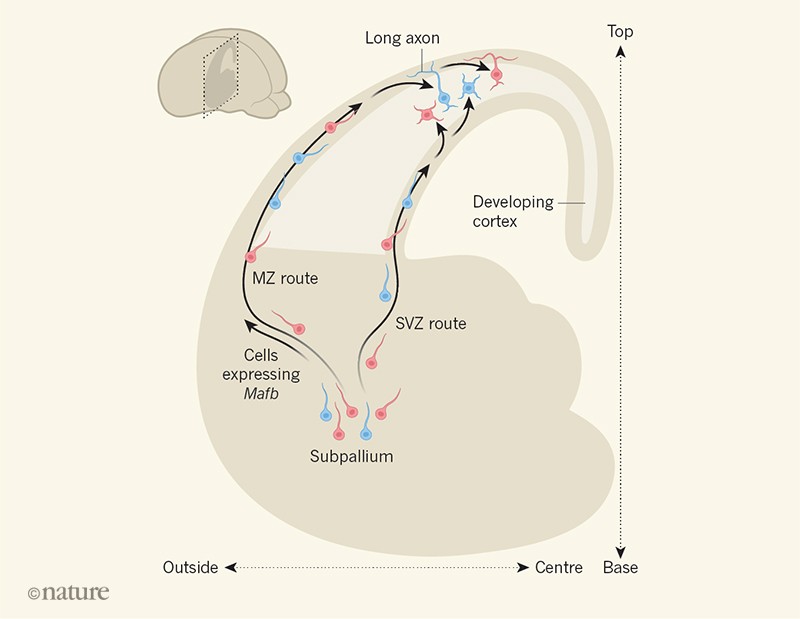
Figura 1 | Las señales intrínsecas y ambientales rigen el desarrollo de interneuronas. Las células llamadas interneuronas se vuelven progresivamente más diversas a medida que maduran. Las interneuronas generadas en el subpallium del cerebro del ratón se pueden dividir en un grupo que se convertirá en células maduras que expresan la proteína somatostatina (roja) y otra que expresará la proteína parvalbúmina (azul). Las neuronas de cada grupo migran a la corteza a través de una de dos rutas: a lo largo de la zona marginal (MZ) por encima de la corteza en desarrollo o a lo largo de la zona subventricular (SVZ) debajo de ella. Lim et al. informaron que una señal intrínseca - expresión del gen Mafb- lleva a las células a migrar a través de la MZ, y a desarrollar largas proyecciones axónicas cuando se mueven hacia la corteza, mientras que las células que migran a través de la ZSV desarrollan axones locales cortos. Sin embargo, la ruta de migración y el desarrollo de las proyecciones también dependen de señales ambientales, que a menudo implican actividad neuronal (no se muestra). La orientación del cerebro está indicada en el recuadro y con flechas punteadas. (Nature 560 , 39-40 (2018). doi: 10.1038 / d41586-018-05737-2)
A continuación, los autores investigaron las consecuencias de eliminar genes que se expresan en los precursores de las interneuronas-somatostatina que migran a través de la zona marginal, pero no en aquellos que pasan a través de la zona subventricular. El grupo descubrió que la eliminación de uno de estos genes, Mafb, en estas células produce una disminución de aproximadamente 20% en la fracción de precursores de las interneuronas-somatostatina que migran a través de la zona marginal. Además, aquellas neuronas que no pudieron migrar a través de la zona marginal carecían de sus proyecciones translaminares características. Finalmente, Lim et al. aislaron las interneuronas migratorias de ambas rutas y las trasplantaron de nuevo al comienzo de la ruta de migración en una porción de cerebro cultivada. Poco más del 60% de las células de la zona marginal volvieron a la misma ruta de migración, lo que sugiere que las diferencias intrínsecas entre las neuronas influyen en sus rutas de migración celular.
Tomando sus resultados en conjunto, Lim y sus colegas concluyen que, al principio del desarrollo, los factores genéticos determinan en qué tipo de interneurona se convertirá una célula y dirigen a la célula por el camino migratorio apropiado. Sin embargo, también hay evidencia del trabajo actual sobre los efectos ambientales sobre el desarrollo interneuronal.
En primer lugar, los efectos de la ruta de migración en la ramificación axonal parecen ser en gran medida independientes del cebado genético, porque los precursores de las neuronas que expresan somatostatina y parvalbúmina se ven afectados de manera similar. En segundo lugar, aunque las células deficientes en Mafb que no migran a través de la zona marginal carecen de sus proyecciones translaminares, conservan otras propiedades características de las células que siguen la ruta de la zona marginal. En tercer lugar, casi el 40% de las interneuronas trasplantadas en cortes cerebrales de la zona marginal eligieron una ruta migratoria diferente en la segunda vuelta. Por lo tanto, es probable que los procesos estocásticos sean una parte importante de la distribución de interneuronas entre las rutas de migración. Parece que está surgiendo un equilibrio entre los aspectos predeterminados y ambientalmente especificados del desarrollo interneuronal.
Una explicación de cómo funcionaría tal equilibrio implica la expresión de genes como Mafb en interneuronas recién nacidas que actúan como una virtual 'tabla de consulta'. En este escenario, la expresión de señales intrínsecas podría sesgar la respuesta de las neuronas en desarrollo a las señales ambientales posteriores. Como tal, la combinación de la expresión génica temprana junto con señales ambientales posteriores podría determinar conjuntamente la identidad y conectividad final de las células.
La determinación de las influencias de las señales ambientales en poblaciones particulares de interneuronas requiere la capacidad de dirigirse selectivamente a esas poblaciones. La identificación de este año de los genes expresados al principio del desarrollo, que son específicos de poblaciones interneuronales particulares, promete proporcionar una forma de sondear las contribuciones de señales intrínsecas tempranas y posteriores del entorno en subclases interneuronales particulares. Ciertamente, el presente documento proporciona una fuerte evidencia de cómo los dos aspectos del desarrollo están vinculados.
Nature 560 , 39-40 (2018)
doi: 10.1038 / d41586-018-05737-2



Comentarios